
Коллектив российских и зарубежных ученых описал новую супергруппу протистов, названную Provora — «прожорливые протисты». Проворы — мелкие (3–15 мкм) жгутиконосцы, питающиеся другими протистами. К проворам относятся две клады — Nebulidia и Nibbleridia, сильно различающиеся по ультраструктуре, способу питания и геному. Эти протисты оказались очень широко распространенными. Их удалось обнаружить в разнообразных морских биотопах от полярных широт до тропиков, а также в нескольких пресноводных местообитаниях, однако в почве они не найдены. Видимо, эти хищные протисты, как и хищные животные, относительно немногочисленны по сравнению с их жертвами, а потому их не обнаруживали раньше в ходе метагеномных исследований.
К успеху часто приводит сотрудничество ученых разных специальностей, и открытие новой группы протистов — как раз такой случай. В коллектив авторов новой работы входят биоинформатики из разных учреждений и ведущие российские протистологи — Д. В. Тихоненков и ныне покойный А. П. Мыльников из Института биологии внутренних вод РАН, а также С. А. Карпов из Зоологического института РАН. (Хотя еще вопрос, насколько протистология и биоинформатика сейчас — разные специальности? Может быть, залог успеха в том, что для многих продвинутых ученых они уже слились воедино...)
Всякому, кто внимательно рассматривал на среднем увеличении микроскопа пресноводные пробы, наверняка знакома такая картинка: наряду с крупными, радующими глаз инфузориями и коловратками в поле зрения в массе движутся мелкие шарики, капельки и запятые. Если глядящий в микроскоп школьник или студент замечает их и задает вопрос: «А это что за мелочь?», приходится отвечать: «Для бактерий вроде крупноваты, хотя кто его знает... Скорее всего, это какие-то мелкие гетеротрофные жгутиконосцы. Точнее определить их не получится.» И вот за последние годы постепенно выясняется, что среди этих невнятных жгутиконосцев, которые и для специалистов-то все часто на одно лицо, скрывается едва ли не львиная доля разнообразия эукариот — точнее, базальных «веточек» их эволюционного древа.
Выделить и описать новую «супергруппу» удалось благодаря сочетанию молекулярных методов с классическими — культивированием отдельных линий и изучением их морфологии. Авторы отмечают, что методы культивирования возрождаются в последние десятилетия, поскольку они оказались незаменимыми в выявлении разнообразия эукариот. Ведь именно среди эукариот распространена такая стратегия питания, как хищничество. А хищники в природных сообществах обычно довольно малочисленны по сравнению с жертвами, и при метагеномных исследованиях их ДНК часто не привлекает внимания — она рассматривается как «сиротская» (то есть неизвестно чья) или вовсе исключается из анализа. Без специальных методов филогеномики особенно сложно бывает понять, кому принадлежит выделенная ДНК, если речь идет о достаточно древней и сильно дивергировавшей от других эукариот группе. Вот почему методы высокопроизводительного секвенирования недостаточны для выявления реального разнообразия хищных эукариот — их приходится дополнять культивированием (что, вообще-то, желательно и для других групп организмов). И такая тактика уже не в первый раз позволяет выявить новые интересные группы. Так был открыт, например, хищный родственник красных водорослей Rhodelphis (см. Открыт хищный родственник красных водорослей, «Элементы», 30.07.2019), а также новая групп хищных альвеолят — акавомонады (см. D. V. Tikhonenkov et al., 2014. Description of Colponema vietnamica sp. n. and Acavomonas peruviana n. gen. n. sp., Two New Alveolate Phyla (Colponemidia nom. nov. and Acavomonidia nom. nov.) and Their Contributions to Reconstructing the Ancestral State of Alveolates and Eukaryotes).
Залог успеха изучения хищных протистов — создание подходящих культуральных условий. В обсуждаемой работе в пробу сначала добавляли бактерий Pseudomonas fluorescens, чтобы стимулировать размножение питающихся ими мелких жгутиконосцев. Это, в свою очередь, стимулировало размножение провор. Затем их клетки вылавливали микропипеткой (что не так-то просто сделать из-за их мелких размеров и высокой скорости плавания) и культивировали в морской воде, добавляя в качестве пищи мелких морских жгутиконосцев-бодонид (Bodonida) Procryptobia sorokini.
Живые проворы (с латыни их название можно перевести как «прожорливые протисты») были обнаружены в пробах из самых разных морских местообитаний — с поверхности кораллов и губок на коралловых рифах в Красном море и возле Кюрасао, из прибрежных донных осадков Черного и Красного морей, из толщи воды с глубин от 11 до 220 м северо-восточной Пацифики и Северного Ледовитого океана. Еще более широкое их распространение показал анализ собранных ранее данных по 18S рРНК: когда стало ясно, что искать, ДНК провор была обнаружена в пробах из всех океанов, в том числе на мезопелагических глубинах (несколько сотен метров) в толще воды и в донных отложениях абиссали (5000 м). Удалось обнаружить их гены и в двух пресноводных местообитаниях в Канаде — в солоноватом озере и в пресном ручье. А вот в почвенных пробах гены провор отсутствовали.
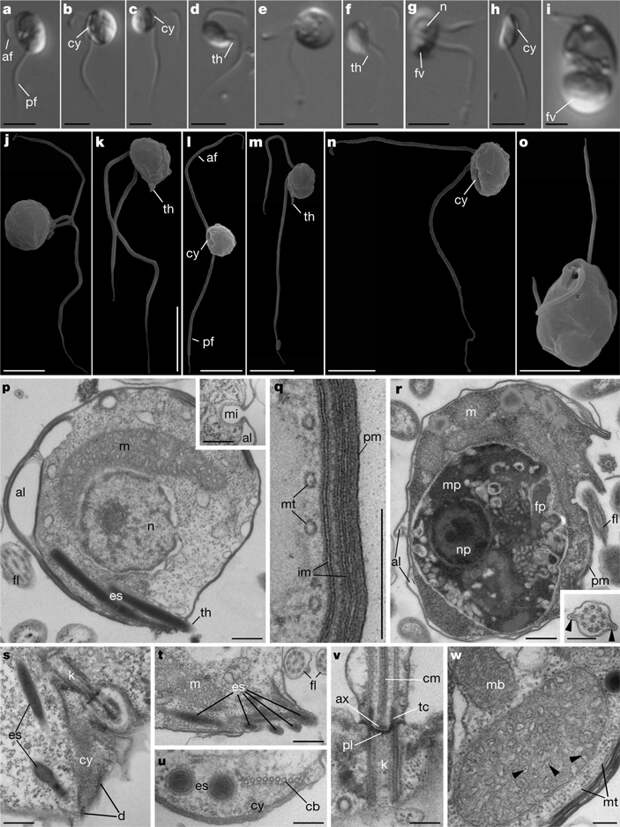
Ультраструктура новых групп, к сожалению, описана в статье не слишком подробно. В общем диагнозе указано только, что это свободноживущие хищные жгутиконосцы с двумя жгутиками, вентральной пищевой бороздкой (так называется цитостом вытянутой формы) и экструсомами. Это связано, в частности, с заметными различиями строения двух групп провор — нибблерид (Nibbleridia) и небулид (Nebulidia).
Из последующих описаний следует, что обе группы имеют гетеродинамные жгутики, выходящие из двух отдельных околожгутиковых карманов. У одного из видов небулид передний жгутик короче (равен длине клетки) и совершает «машущие движения», а задний вдвое длиннее клетки и ундулирует в проксимальной части. Характерно ли это для всех провор, не ясно. Кроме того, все виды имеют сложные клеточные покровы, включающие в себя альвеолы. При этом подчеркивается, что эти признаки встречаются и у многих других групп протистов и не говорят о тесном родстве.
Чтобы детальнее разобраться в строении клеток провор, приходится смотреть более ранние их описания. В деталях нибблериды и небулиды достаточно сильно различаются по морфологии. Небулиды крупнее (10–14 мкм) и имеют овальную форму; у нибблерид длина всего 3–6 мкм, а их «голодные» клетки заметно изогнуты. Передний жгутик небулид несет у основания мастигонемы, а у нибблерид они отсутствуют. У нибблерид жгутики имеют акронему — тонкий концевой участок, у небулид акронемы нет. Задний жгутик нибблерид несет две боковые складки (рис. 1), а задний жгутик небулид — только одну и только в проксимальной части (авторы кое-где называют эту складку «жилкой», хотя ее длина больше диаметра жгутика, и на жилку она мала похожа).
Судя по тексту, эта складка латеральная; но на самом деле, кажется, положение складок жгутиков провор относительно их тела не выяснено. У нибблерид переходная зона жгутиков содержит промежуточный цилиндр, расположенный дистальнее вогнутой поперечной пластинки (рис. 1). Переходная зона небулид в статье не описана, а из более ранних работ вроде бы следует, что поперечных пластинок в промежуточном цилиндре две — более тонкая проксимальная и более толстая дистальная. Кристы митохондрий нибблерид в статье описаны как мешковидные, а в дополнительных материалах к ней (таксономические описания) указано, что в митохондриях есть мешковидные и трубчатые кристы. В просвете крист у этих протистов присутствуют странные нитевидные включения (рис. 1, w). У небулид, по-видимому, в митохондриях есть и трубчатые, и пластинчатые кристы; нитевидные включения в них не описаны.
Покровы клеток небулид, кроме плазмалеммы (в прежних статьях авторы почему-то называли так одну из внутренних мембран, но в обсуждаемой статье это исправлено), включают две или три внутренних мембраны; между ними довольно равномерно расположены мембранные везикулы. Такая оболочка окружает всю клетку, кроме пищевой бороздки. Что из этого гомологично альвеолам других протистов, сказать сложно. Покровы нибблерид в голодном состоянии включают три-четыре слоя внутренних мембран под плазмалеммой, один-два слоя пузырьков (альвеол?) под ними на дорзальной стороне, небольшие впячивания (микропоры) между альвеолами и равномерно расположенные микротрубочки под ними. У «сытых» клеток внутренних слоев мембран нет; предполагается, что они используются для образования крупной пищеварительной вакуоли. Хотя обе группы имеют экструсомы, они разного строения. У нибблерид это 5–6 трихоцист внутри заостренного выроста — рога, расположенного сзади от пищевой бороздки (рис. 1, k, m, t). У небулид экструсомы сидят ближе к переднему концу клетки. У одного их представителя — Nebulomonas — это токсицисты (состоят из двух цилиндров, из которых внутренний, видимо, выдвигается после выстреливания) (рис. 2).

У другой небулиды — Ancoracysta — описан особый тип экструсом, анкорацисты (рис. 3).

На поперечном срезе анкорацисты выглядят как мандарин с семью дольками, а на их переднем конце есть расширенная шапочка, которая на продольном срезе выглядит как лапки якоря. У всех провор экструсомы, вероятно, служат для нападения на добычу, каким-либо способом ее обездвиживая.
Наконец, различается пищевое поведение провор. Обе группы охотятся на жертв, сравнимых по размерам с хищником, и во всех случаях жертва попадает в крупную пищеварительную вакуоль в задней части клетки. При этом небулиды глотают добычу целиком. Нибблериды к этому тоже способны, но у них обнаружен и второй способ питания (см. видео ниже) — они умеют откусывать куски от добычи! Возможно, эту уникальную для протистов способность обеспечивают «зубы» — электронноплотные выросты на краях цитостома.
Но позвольте — о каких «более ранних работах» идет речь, если проворы — новая группа протистов? На самом деле в статье впервые описаны только нибблериды — четыре вида рода Nibbleromonas и один вид рода Ubysseya (назван он так в честь студенческой газеты Ubyssey Университета Британской Колумбии (UBC), где работает последний автор статьи, Patrick J. Keeling). Два известных вида небулид были описаны ранее. Первый, Nebulomonas marisrubri, был найден в интерстициали песчаного пляжа в Красном море и описан в 2009 году (А. П. Мыльников, Д. В. Тихоненков, 2009. Новый альвеолятный хищный жгутиконосец Сolponema marisrubri (Сolponemida, Аlveolata) из Красного моря). Второй вид, Ancoracysta twista, был собран с поверхности коралла-мозговика в тропическом морском аквариуме и описан в 2017 году (J. Janouškovec et al., 2017. A New Lineage of Eukaryotes Illuminates Early Mitochondrial Genome Reduction; про это открытие «Элементы» уже рассказывали в новости Новооткрытый жгутиконосец проливает свет на раннюю эволюцию эукариот). При этом Nebulomonas marisrubri была описана Тихоненко и Мыльниковым как Colponema marisrubri, то есть как представитель колпонемид — таксона в составе альвеолят, выделенного ими же (с соавторами) в отдельный тип в 2014 году. Ancoracysta twista изначально была описана как загадочный вид неизвестного таксономического положения.
Кажется, первым догадался о родстве C. marisrubri и Ancoracysta twista выдающийся протистолог Томас Кавалье-Смит. В своей статье Multigene phylogeny and cell evolution of chromist infrakingdom Rhizaria: contrasting cell organisation of sister phyla Cercozoa and Retaria, опубликованной в 2018 году совместно с коллегами по Оксфорду, он указал на многочисленные сходные ультраструктурные признаки этих двух видов и на отличия Colponema marisrubri от других колпонемид, предложив включить ее в род Ancoracysta. Тут, правда, проявился некоторый «экстремизм»: утверждалось, что токсицисты C. marisrubri и анкорацисты Ancoracysta чуть ли не идентичны. Судя по поперечным срезам, это явно не так, и первооткрыватели с помещением этих таксонов в один род не огласились.
Из всех этих перипетий следует, что анализ ультраструктуры, как и любых морфологических признаков, увы, слишком часто приводит к субъективным трактовкам и выводам. В статье с описанием провор, например, как «вероятно, древние, напоминающие предковое состояние для нескольких крупных супергрупп эукариот» трактуются такие признаки нибблерид, как две продольные складки жгутика (встречаются также у малавимонад, некоторых метамонад и дискобид) и нитчатые структуры крист митохондрий (есть у некоторых золотистых и желто-зеленых водорослей). На деле, конечно, такое «мозаичное» распределение признака среди неродственных групп ничего не говорит о его древности, с тем же успехом это может оказаться и результатом конвергенции (см. обсуждение этой проблемы в статье J. Lukeš et al., 2009 Cascades of convergent evolution: The corresponding evolutionary histories of euglenozoans and dinoflagellates).
К появлению на свет провор как супергруппы научная общественность была подготовлена. В статье 2020 года Predatory flagellates — the new recently discovered deep branches of the eukaryotic tree and their evolutionary and ecological significance на их существование намекает Д. В. Тихоненков (тут, правда, Ancoracysta отнесена еще не к проворам; она фигурирует как сестринская группа Haptista (гаптофитовые водоросли + центрохелидные солнечники), а будущие нибблериды — как сестринская группа Haptista + TSAR). Теперь родственники счастливо объединились, но до сих пор осталось не до конца ясным, от кого же они отделились. Генетические данные дают разные результаты в зависимости от метода анализа. Байесовский вывод (Bayesian inference) дает дерево, на котором проворы — сестринская группа Haptista + TSAR. Метод максимального правдоподобия показывает, что проворы — сестринская группа гемимастигофор, еще одной «загадочной группы» протистов. Зато теперь мы точно знаем, что небулиды и нибблериды — хоть и очень давно разошедшиеся, но относящиеся к одному монофилетическому таксону клады.
В целом геномы провор оказались достаточно крупными для протистов — в них насчитывается примерно от 11 000 до 24 000 генных семейств, причем общих для нибблерид и небулид всего около 20–25%. Несмотря на предполагаемый небольшой размер популяций провор, в их геномах не обнаружено признаков ускоренной эволюции. По-видимому, у предков провор не было пластид, так как никакие «пластидные» гены у них не найдены. Обогащены их геномы генами, связанными с кальциевой сигнализацией (в том числе с разными кальций-зависимыми каналами). Обогащение это произошло независимо у небулид и нибблерид; возможно, эти гены связаны с регуляцией клеточной подвижности и других форм поведения провор. Еще сильнее геномы обогащены белками (точнее, белковыми доменами), которые у других эукариот участвуют в образовании мембраноатакующих комплексов или играют роль перфоринов. И опять-таки, увеличение числа генов этих семейств (вероятно, связанных с работой экструсом) произошло независимо в разных линиях провор.
Митохондриальные геномы всех провор тоже богаты генами — они одни из самых больших среди всех существующих. У всех провор митохондрии содержат консервативный набор из 51 белкового гена, и в целом в их митохондриальном геноме есть лишь небольшие вариации. Вариации состоят в наличии/отсутствии нескольких генов рибосомальных белков и нескольких генов, кодирующих тРНК, а также бактериального типа rnpB (кодирует каталитическую РНК, участвующую в созревании тРНК). В большинстве случаев «недостающие» гены, видимо, были не утрачены, а перенесены в ядро — их продукты обнаруживаются в транскриптомах. Все митохондриальные геномы Provora кодируют компоненты системы созревания цитохрома c I типа (ccmA, ccmB, ccmC and ccmF), которые они унаследовали от бактериального предка митохондрий (чтобы цитохром c работал, нужно к белку апоцитохрому присоединить гем; этим и занимаются системы созревания цитохромов). У небулид, кроме системы типа I, есть система созревания цитохрома c III типа — голоцитохром c синтаза (HCCS), закодированная в ядерном геноме. Эта система заменила систему созревания I типа у большинства эукариот, а у остальных протистов, сохранивших систему типа I, она отсутствует (подробнее см. новость Новооткрытый жгутиконосец проливает свет на раннюю эволюцию эукариот, «Элементы», 17.01.2018).
Теперь мы знаем, что наличие двух систем одновременно — не уникальная черта Ancoracysta. Раз и у N. marisrubri, и у A. twista присутствуют две системы созревания цитохромов — значит, обе системы сосуществовали в течение протяженного времени в процессе эволюции. Изучение митохондрий провор, как надеются авторы статьи, может дать уникальную информацию об эволюции системы созревания цитохрома c у эукариот.
Хорошо бы, конечно, постепенно разобраться и в генетических основах ультраструктурных различий протистов (хотя бы в том, чем отличаются протеомы токсицист и анкорацист, или в том, какие гены отвечают за наличие двух складок на жгутиках у неродственных групп). Но это пока дело будущего.
Поиски новых групп протистов — не менее интересное занятие. И особенно интересны базальные веточки эволюционного древа — тоннель в стене, еще недавно наглухо отделявшей прокариот от эукариот, можно ведь надеяться пробить не только «со стороны» архей (см. новость У асгардархей обнаружен сложный актиновый цитоскелет, «Элементы», 09.01.2023).
Источник: Denis V. Tikhonenkov, Kirill V. Mikhailov, Ryan M. R. Gawryluk, Artem O. Belyaev, Varsha Mathur, Sergey A. Karpov, Dmitry G. Zagumyonnyi, Anastasia S. Borodina, Kristina I. Prokina, Alexander P. Mylnikov, Vladimir V. Aleoshin & Patrick J. Keeling. Microbial predators form a new supergroup of eukaryotes // Nature. 2022. DOI: 10.1038/s41586-022-05511-5.
Сергей Глаголев
Свежие комментарии