
Эволюция всех сложных форм земной жизни стала возможной благодаря тому, что предки современных эукариот приобрели внутриклеточных симбионтов, впоследствии ставших органеллами — митохондриями и хлоропластами. Превращение бактериального симбионта в органеллу — очень редкое эволюционное событие. До сих пор было известно всего три таких случая (третий — независимо приобретенная фотосинтезирующая органелла амёбы Paulinella). Четвертый случай удалось обнаружить в ходе изучения морской одноклеточной водоросли Braarudosphaera bigelowii. Как выяснилось, азотфиксирующий симбионт браарудосферы, считавшийся симбиотической цианобактерией, настолько глубоко интегрирован в хозяйскую клетку, что его правильнее считать не симбионтом, а органеллой нового типа — нитропластом. Почти половина белков, работающих в нитропласте, кодируются ядерным геномом водоросли и импортируются в нитропласт специальными транспортными системами, а деление нитропласта аккуратно синхронизировано с жизненным циклом клетки-хозяина: нитропласт делится после митохондрии, но до хлоропластов и ядра. Открытие проливает свет на ранние этапы эволюции органелл и может оказаться важным для решения биоинженерных задач.
Эволюционный успех эукариот неразрыно связан с наличием у них органелл бактериального происхождения — митохондрий и хлоропластов. Благодаря митохондриям (потомкам симбиотической альфа-протеобактерии) эукариоты способны к кислородному дыханию, а хлоропласты (потомки симбиотической цианобактерии) подарили водорослям и высшим растениям способность к оскигенному фотосинтезу.
Эукариоты легко заводят разнообразных бактериальных симбионтов, в том числе внутриклеточных. Но вот превратить такого симбионта в интегральную часть клетки — органеллу, по-видимому, очень сложная задача для эволюции. Считается, что митохондрии всех эукариот имеют единое просхождение, то есть являются результатом одного-единственного события «первичного эндосимбиоза», произошедшего более двух миллиардов лет назад. Та же ситуация и с хлоропластами: все они происходят от одного вида цианобактерий, одомашненного общим предком архепластид (см. Archaeplastida) 2,0–1,5 млрд лет назад.
Впоследствии было еще много событий «вторичного» и «третичного» эндосимбиоза, когда нефотосинтезирующие эукариоты приобретали в качестве внутриклеточных симбионтов (и впоследствии превращали в органеллы) уже не цианобактерий, а одноклеточные эукариотические водоросли с готовыми хлоропластами (см. Геномы одноклеточных водорослей проливают свет на эволюцию фотосинтезирующих эукариот, «Элементы», 07.12.2012). По-видимому, вторичные и третичные эндосимбиозы по каким-то причинам являются более простыми (вероятными) эволюционными событиями, чем первичный эндосимбиоз. Единственное исключение — амёба Paulinella, которая сравнительно недавно (около 120 млн лет назад, то есть в раннем мелу) независимо обзавелась фотосинтезирующей органеллой — «хроматофором». Хроматофор происходит от симбиотической цианобактерии, неродственной предку хлоропластов.
Органелла отличается от обычной эндосимбиотической бактерии двумя важнейшими особенностями. Во-первых, она полностью интегрирована в архитектуру хозяйской клетки, а ее размножение синхронизировано с жизненным циклом хозяина. Второе и главное: жизнедеятельность органеллы определяется и контролируется многочисленными белками, закодированными в ядерном геноме хозяина. Эти белки транспортируются в органеллу специальными транспортными системами. Обычно у таких белков на конце есть специальный сигнальный участок — транзитный пептид (transit peptide), по которому клетка определяет, что белок нужно доставить в органеллу. Считается, что благодаря такому централизованному контролю сотрудничество клетки-хозяина с органеллой в итоге оказывается гораздо более эффективным и надежным, чем с обычной симбиотической бактерией.
Еще один характерный признак древних органелл (митохондрий и пластид) состоит в том, что многие гены, изначально находившиеся в геноме симбионта, перенесены в ядерный геном. Однако у паулинеллы в ядерном геноме почти нет генов, проиcходящих из генома ее молодой органеллы. Возможно, массовый перенос генов в ядерный геном, сопряженный с экстремальной редукцией генома органеллы, происходит на более поздних этапах, когда система транспорта белков в органеллу уже хорошо отлажена. А поначалу в органеллу транспортируются в основном белки эукариотического происхождения или заимствованные у кого-то еще путем горизонтального переноса (см. Stephens et al., 2021. Why is primary endosymbiosis so rare?; в этой статье представлена гипотетическая модель поэтапной эволюции органелл, основанная на изучении паулинеллы и ее хроматофора; новые данные хорошо согласуются с этой гипотезой).
Помимо кислородного дыхания и фотосинтеза есть еще один крайне важный и полезный биохимический процесс, освоенный прокариотами, а эукариотам доступный только через сотрудничество с бактериальными симбионтами. Это азотфиксация — перевод азота из практически недоступной для живых существ формы N2 в легко усвояемый аммиак или аммоний (NH3, NH4+). Многие эукариоты, в том числе высшие растения, водоросли, грибы и даже некоторые животные, неоднократно вступали в сложные, порой очень тонко отлаженные и эффективные симбиозы с азотфиксирующими бактериями (см. ссылки в конце новости). Но удалось ли хоть кому-то из-них превратить азотфиксирующего симбионта в полноценную органеллу — нитропласт?
Исследование американских, японских и тайваньских биологов, результаты которого опубликованы недано в журнале Science, показывает, что как минимум одна одноклеточная водоросль сумела это сделать.
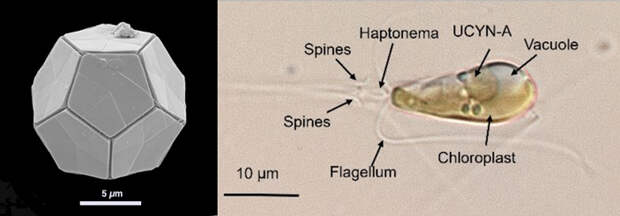
Речь идет о широко распространенной в морях и океанах гаптофитовой водоросли Braarudosphaera bigelowii. Эта водоросль генетически довольно разнообразна и по сути представляет собой не единый вид, а группу родственных видов-близнецов. Браарудосфера относится к кокколитофоридам — гаптофитовым водорослям, образующим на поверхности клетки известковые пластинки разнообразной формы — кокколиты (рис. 1). Кокколитофориды играют важную роль в круговороте углерода (см. Фитопланктон реагирует на рост концентрации CO2 не так, как ожидалось, «Элементы», 21.04.2008) и, может быть, отчасти защищают биосферу от пагубного влияния резких скачков концентрации CO2 в атмосфере (см. Связь массовых вымираний с вулканизмом получила новое подтверждение, «Элементы», 19.09.2011).
Кокколиты отлично сохраняются в ископаемом состоянии. Поэтому мы знаем, что браарудосфера живет в океанах с начала позднемеловой эпохи (100 млн лет назад), а ее близкая родня процветала уже в раннем мелу. Обычно браарудосфера была сравнительно редкой водорослью, но она хорошо переживала кризисы и становилась массовой после крупных вымираний, когда разнообразие фитопланктона падало (в том числе в датском веке, после великого мел-палеогенового вымирания).
Не исключено, что такая жизнестойкость отчасти связана с тем, что браарудосфера способна к азотфиксации. Для этого у нее есть специальный симбионт — цианобактерия, известная под условным названием Candidatus Atelocyanobacterium thalassa (UCYN-A). Ученые давно интересуются этим симбиозом, но толком изучить его долго не удавалось, потому что культивировать браарудосферу в лаборатории научились лишь недавно (Suzuki et al., 2021. Unstable Relationship Between Braarudosphaera bigelowii (= Chrysochromulina parkeae) and Its Nitrogen-Fixing Endosymbiont).
Авторам обсуждаемой статьи удалось показать, что азотфиксирующий симбионт UCYN-A заслуживает высокого звания органеллы не в меньшей степени, чем хроматофор паулинеллы.
При помощи мягкой рентгеновской томографии (SXT, soft-x-ray-tomography) ученые построили детальные трехмерные реконструкции клеток в разные периоды суточного цикла (днем браарудосфера занимается фотосинтезом и азотфиксацией, а ночью размножается делением). В неделящейся клетке браарудодосферы есть два хлоропласта (это «вторичные», четырехмембранные хлоропласты, происходящие от симбиотической красной водоросли, как у всех гаптофитов), одна большая разветвленная митохондрия, одно ядро и один крупный (занимающий около 9% объема клетки) шарообразный азотфиксирующий симбионт UCYN-A. Он всегда находится у заднего конца клетки, и к нему плотно прилегает митохондрия. Это имеет смысл, потому что азотфиксация — крайне энергозатратный процесс, а митохондрия — это «энергетическая станция» клетки. Она производит АТФ, необходимый для азотфиксации.
Когда приходит пора делиться, первой делится митохондрия, за ней UCYN-A, и лишь после этого наступает черед ядра и пластид (рис. 2).

Таким образом, налицо признаки глубокой интеграции UCYN-A в архитектуру хозяйской клетки: фиксированное положение в задней части клетки, обязательный контакт с митохондрией, деление в строго определенный момент — после митохондрии, но до хлоропластов и ядра.
Это серьезный аргумент в пользу того, что UCYN-A — не просто симбионт, а органелла. Второй аргумент, еще более весомый, дали результаты протеомного анализа. Это трудоемкая задача, ведь, чтобы понять, какие белки транспортируются из цитоплазмы в UCYN-A, одним секвенированием геномов и транскриптомов не обойтись: нужно работать с самими белками. Авторы сначала научились аккуратно выделять UCYN-A из разрушенных клеток при помощи центрифугирования. Белки, выделенные по отдельности из очищенных UCYN-A и из целых клеток браарудосферы, разрезались ферментами на короткие кусочки (пептиды), массу которых определяли при помощи масс-спектрометрии. Поскольку и ядерный геном браарудосферы, и геномы всех ее органелл прочтены, и известно, какие белки в них закодированы, а также известно, на какие именно кусочки должен был разрезаться каждый из них, полученные результаты позволили идентифицировать в пробах много конкретных белков (хотя, конечно, далеко не все).
В общей сложности на этом этапе было идентифицировано 1804 белка, из которых 1156 закодированы в ядерном геноме и 609 — в геноме UCYN-A. Почти все белки, закодированные в UCYN-A, там же и локализуются. Но вот из белков, закодированных в ядре, заметная часть (368) присутствует в UCYN-A в значимо большем количестве, чем в целых клетках. Стало быть, многие закодированные в ядре белки импортируются в UCYN-A.
Посмотрев на геномные последовательности этих 368 белков, ученые обнаружили, что большинство из них значительно длиннее, чем соответствующие (ортологичные) белки других водорослей. Разница в длине объясняется наличием у белков, импортируемых в UCYN-A, дополнительного участка на C-конце белка. У разных белков, импортируемых в UCYN-A, этот участок похож по своей аминокислотной последовательности, но при этом он не имеет никакого сходства с другими известными белками. Судя по всему, это эволюционное новообразование — «транзитный пептид», сигнализирующий о том, что данный белок должен быть импортирован в UCYN-A. Следовательно, у браарудосферы есть специализированная система транспорта белков в UCYN-A, ориентирующаяся в своей работе на особый транзитный пептид — так же, как и транспортные системы, доставляющие белки в хроматофор паулинеллы, митохондрии и хлоропласты. Транзитный пептид UCYN-A получил название uTP (UCYN-A transit peptide).
Оказалось, что uTP есть у многих белков, закодированных в ядерном геноме, но не выявленных в ходе протеомного анализа. Скорее всего, эти белки, ускользнувшие от несовершенного протеомного анализа, тоже импортируются в UCYN-A. Объединив протеомные и геномные данные, ученые получили список из 900 импортируемых белков. Это существенная прибавка к 1186 белкам, закодированным в геноме UCYN-A.
Как и в случае с паулинеллой и ее хроматофором, в ядерном геноме браарудосферы нет генов, перенесенных из UCYN-A в ядро. Большинство белков, импортируемых в UCYN-A, имеют эукариотическое происхождение. Это согласуется с вышеупомянутой гипотезой о том, что массовый перенос генов из органеллы в ядро начинается на более поздних этапах эволюции органеллы.
Тем не менее геном UCYN-A сильно упрощен по сравнению с геномами свободноживущих цианобактерий (хотя, конечно, не так сильно, как геномы хлоропластов и митохондрий). В нем отсутствуют многие необходимые свободноживущим цианобактериям гены. Из-за этого разнообразные биохимические пути (в том числе пути синтеза аминокислот, кофакторов и нуклеотидов) оказываются «дырявыми», то есть для их реализации не хватает одного или нескольких ферментов. Во многих случаях эти пробелы компенсируются импортируемыми белками. Есть также случаи «избыточности», когда в UCYN-A есть и свой собственный, цианобактериальный белок с данной функцией, и его импортный аналог, закодированный в ядерном геноме водоросли. По-видимому, упрощение генома UCYN-A идет по схеме «белок начинает импортироваться извне — возникает избыточность — собственный ген теряется».
Обнаруженные факты (структурная интергация, синхронизация деления, импорт хозяйских белков) показывают, что UCYN-A можно по праву назвать органеллой нового типа — нитропластом. Причем это молодая органелла, находящаяся примерно на той же стадии своего эволюционного пути (от вольной бактерии к старой органелле вроде хлоропласта), что и хроматофор паулинеллы (см. таблицу).
Сравнение геномов цианбоактерий и произошедших от них органелл (из дополнительных материалов к обсуждаемой статье, с небольшой правкой на основе других источников)
| Цианобактерия или органелла | Длительность эндосимбиоза (млн лет) | Размер генома (млн пар оснований) | Белки, закодированные в собственном геноме | Импортируемые белки, закодированные в ядерном геноме |
| Хлоропласт Arabidopsis | 1500–2000 | 0,1–0,2 | 61–139 | 3561 |
| Хроматофор Paulinella | 120 | 1,0 | 867 | 433 |
| Нитропласт Braarudosphaera (UCYN-A) | 100 | 1,5 | 1186 | 900 |
| Цианобактерия Cyanothece, родственная UCYN-A | — | 4,8–5,5 | 4444–5304 | — |

Азотфиксирующие эндосимбионты цианобактериального происхождения встречаются не только у гаптофитовых, но и у диатомовых водорослей, причем как морских, так и пресноводных. Азотфиксирующие симбионты морских диатомей происходят от цианобактерий, родственных предкам UCYN-A. Однако эти симбиозы, по-видимому, значительно моложе и соответствуют более ранним этапам пути от бактерии к органелле (Schvarcz et al., 2022. Overlooked and widespread pennate diatom-diazotroph symbioses in the sea). Впрочем, данный вопрос еще недостаточно изучен. Хотя формально эти симбионты не имеют статуса органелл, их всё равно иногда называют «нитропластами» или «диазопластами» (что по смыслу одно и то же).
Эукариотические водоросли с азотфиксирующими симбионтами — не какая-нибудь экзотика, никому не интересная, кроме биологов-эволюционистов. В бедных азотом областях океана они играют важную роль в круговороте азота и вносят заметный вклад в биологическую продуктивность. Их изучение может когда-нибудь в светлом будущем помочь созданию методами биоинженерии растений, способных к азотфиксации.
Источник: Tyler H. Coale, Valentina Loconte, Kendra A. Turk-Kubo, Bieke Vanslembrouck, Wing Kwan Esther Mak, Shunyan Cheung, Axel Ekman, Jian-Hua Chen, Kyoko Hagino, Yoshihito Takano, Tomohiro Nishimura, Masao Adachi, Mark Le Gros, Carolyn Larabell, Jonathan P. Zehr. Nitrogen-fixing organelle in a marine alga // Science. 2024. V. 384. P. 217–222.
См. также об азотфиксирующих симбионтах:
1) Излишками азота от симбиотических бактерий гриб платит термитам за распространение своих спор, «Элементы», 04.09.2021.
2) Н. А. Проворов, О. Ю. Штарк, Е. А. Долгих, 2016. Эволюция азотфиксирующих симбиозов, основанная на миграции бактерий из микоризных грибов и почвы в ткани растений. Синопсис: Как это было: эволюционные пути формирования азотфиксирующих симбиозов.
3) Муравьи используют азотные удобрения для своих угодий, «Элементы», 23.11.2009.
4) Бактерии-симбионты, разлагающие для термитов древесину, еще и связывают для них атмосферный азот, «Элементы», 11.12.2008.
5) Изменение гена, необходимого для симбиоза растений с грибами, привело к формированию симбиоза с азотфиксирующими бактериями, «Элементы», 12.03.2008.
6) Н. А. Проворов, Е. А. Долгих. 2006. Метаболическая интеграция организмов в системах симбиоза. Синопсис: От биохимического сотрудничества — к общему геному.
Свежие комментарии