
Американские биологи провели эксперимент на мышах, целью которого была оценка влияния «случайности» или «везения» на развитие и жизненный успех особей. Сотню генетически идентичных двухнедельных мышат с вживленными под кожу метками поместили на огороженную лужайку с 16 уютными домиками, в каждом из которых находилась регулярно пополняемая кормушка. За перемещениями мышей следили при помощи системы радиочастотной индентификации (RFID) в течение 46 суток. Эксперимент показал, что между генетически идентичными мышами в таких условиях формируются устойчивые поведенческие различия. У самцов эти различия выражены сильнее, чем у самок, и быстрее усиливаются с возрастом. Предположительно это объясняется тем, что в условиях эксперимента самцы, в отличие от самок, активно конкурировали друг с другом за контроль над дефицитными ресурсами (домиками, кормушками, самками). В результате у самцов небольшие случайные различия, возникавшие в начале жизни, в дальнейшем нарастали, как снежный ком. То есть особи, которым поначалу повезло оказаться в более выигрышном положении, в дальнейшем усиливали свое преимущество, а неудачники оттеснялись все дальше на задворки социальной жизни. Это явление в социальных науках известно под названием «эффект Матфея». Результаты согласуются с популярной идеей о том, что различия в жизненном успехе нередко зависят не столько от способностей и заслуг индивида, сколько от слепого случая.
В научной литературе, как биологической, так и социологической, активно обсуждается идея о том, что различия между индивидами по степени жизненного успеха (будь то число оставленных потомков, социальный статус или богатство, если речь идет о людях) определяются не только (и даже не столько) генетическими особенностями индивида и какими-то фиксированными, измеряемыми параметрами среды, в которой присходило его развитие, но и тем, что в просторечии называют «везением», «удачей» (luck) или, чуть более наукообразно, «случайностью» (contingency). Имеются в виду разные непредсказуемые события и стечения обстоятельств, на которые нельзя повлиять и которые никак не зависят от способностей, талантов и заслуг индивида.
Например, можно представить себе двух абсолютно одинаковых хищников, отправившихся на охоту в одном и том же лесу и действующих по одному и тому же алгоритму. Первому повезло, он наткнулся на добычу и пообедал, а второй не наткнулся и остался голодным. Потом эти хищники встретились, подрались за лучший участок, и сытый прогнал голодного в менее благоприятное для охоты место. В результате «везунчик» в последующие дни будет лучше питаться, и его физическое превосходство над соперником будет расти. Этот простой пример иллюстрирует сразу два важных соображения. Во-первых, последствия случайного везения, поначалу незначительные, могут в дальнейшем нарастать, как снежный ком. В социологии это называют «эффектом Матфея» («имеющему дастся и приумножится, а у неимеющего отнимется и то, что имеет»). Во-вторых, ключевую роль в усилении этих последствий может играть конкуренция, в результате которой индивид получает тот или иной статус в иерархии и занимает ту или иную социальную нишу. В нашем примере важно, что хищники были территориальными и конкурировали за лучший участок. В противном случае начальное везение вряд ли привело бы к далеко идущим последствиям: просто в другой раз повезло бы второму хищнику, и эффекты случайностей сгладили бы друг друга.
В теории эти рассуждения выглядят правдоподобно, но без экспериментальных подтверждений их цена невелика. Однако экспериментально изучать влияние удачи на жизненный успех довольно трудно. Ведь для этого нужно не только проследить развитие признаков, связанных с успехом, в течение жизни особей, но и как-то выровнять генетику подопытных индивидов и условия среды, в которой они развиваются.
Именно это попытались сделать биологи из Корнеллского университета (США) в любопытном эксперименте на генетически идентичных мышах из стандартной лабораторной линии C57BL/6J, результаты которого опубликованы в журнале Science.
На экспериментальную площадку размером 15 на 38 метров с 16 комфортными мышиными домиками, в каждом из которых находилась неиссякающая кормушка (рис. 1), поместили 16 самок с выводками. Выводки различались по возрасту не более, чем на 48 часов. Средний возраст мышат в момент помещения на площадку составлял 14 суток. В общей сложности в выводках было 104 мышонка, из которых до зрелости дожили 90 (причины смерти остальных авторы не установили).
По мнению исследователей, условия на площадке были «экологически релевантными», то есть довольно-таки естественными для домовых мышей, и этим эксперимент выгодно отличается от прежних попыток такого рода, проводившихся в лаборатории.
За два дня до переселения на площадку, то есть примерно в 12-дневном возрасте, каждому мышонку вживили под кожу метку для радиочастотной идентификации (RFID). Эти метки и антенны, установленные в каждом домике, позволяли регистрировать перемещения животных между домиками. Наблюдения продолжались 46 суток. За это время мышата стали взрослыми, но еще не успели обзавестись собственным потомством.
Важно, что в подобных условиях, как было показано ранее, самцы активно конкурируют друг с другом за контроль над дефицитными ресурсами (домиками и находящимися в них самками и кормушками), в то время как самки перемещаются между домиками свободно и друг с другом практически не конкурируют. Поэтому можно было ожидать, что конкуренция будет усиливать случайные различия между самцами, но не между самками. Результаты подтвердили эти ожидания.
На основе данных RFID для каждого индивида каждую ночь отслеживали 17 показателей (поведенческих признаков), отражающих перемещения мышей и их социальные взаимоотношения. Примеры показателей: число домиков, посещенных за ночь (диапазон 0–16, среднее 6,0); число переходов из домика в домик (0–141, в среднем 16); число переходов из одной группы домиков (neighborhood на рис. 1) в другую (0–26, 2,9); число встреченных самцов (то есть самцов, с которыми данная особь находилась одновременно в одном домике или рядом с ним); число встреченных самок; доля времени, проведенного в одиночестве; доля времени, проведенного вместе хотя бы с одним представителем противоположного пола и т. д. (полный список показателей приведен в дополнительных материалах к статье).
Анализ собранных данных показал, что между генетически идентичными мышами в ходе эксперимента сформировались устойчивые поведенческие различия. Устойчивость проявляется в том, что по данным о поведении мыши в один период времени (например, в возрасте 20–30 дней), можно предсказать, как она будет вести себя в другой период, например, в 30–40 дней. Скажем, если она часто перебегала из домика в домик в 30-дневном возрасте, то, скорее всего, будет часто перебегать и в 40-дневном. Это справедливо для обоих полов и для всех 17 признаков, но в разной степени. Поведенческая индивидуальность особи не зависела от того, к какому из 16 выводков она принадлежала и в каком домике или группе домиков она жила в начале эксперимента. Это согласуется с идеей об изначально «случайной» природе поведенческих различий.
Сложная статистическая обработка данных позволила показать, что у самцов устойчивые поведенческие различия были выражены сильнее, чем у самок. Кроме того, поведенческая индивидуальность у самцов быстрее формировалась в ходе развития (рис. 2).
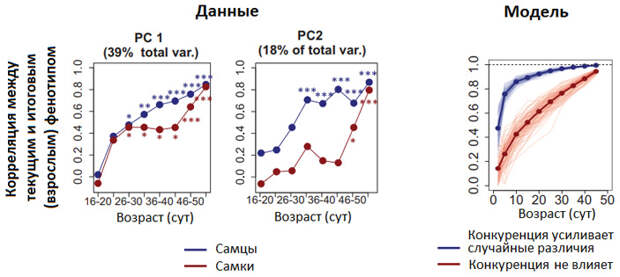
Эти результаты согласуются с идеей о том, что у самцов (но не у самок) случайные поведенческие различия, возникшие в начале жизни благодаря «везению», усиливались с возрастом из-за конкуренции. Например, кому-то из самцов в первые дни самостоятельной жизни (мыши этой линии обычно прекращают питаться материнским молоком в трехнедельном возрасте) случайно повезло захватить контроль над большим количеством домиков, он поверил в себя, лучше питался, испытывал меньше стресса и в результате действительно стал сильнее и конкурентоспособнее других самцов.
Для проверки идеи о том, что поведенческие различия между самцами связаны именно с конкуренцией, авторы рассмотрели динамику показателя, который они назвали «индексом успешности в конкуренции за ресурсы» (Resource Competition Score, RCS). Этот показатель вычислялся для каждого самца и для каждой ночи как сумма долей «мужских» RFID-индентификаций, относящися к данному самцу, во всех домиках, где он побывал. Например, если самец побывал в двух домиках, и в первом из них на его счет приходится 90% всех RFID-идентификаций самцов, а во втором — 40%, то RCS для данного самца в эту ночь будет равен 0,9 + 0,4 = 1,3. По мнению авторов, которое они обосновывают косвенными статистическими аргументами, RCS отражает именно доминантность самца, то есть его успешность в конкуренции с другими самцами (а, например, не склонность малодушно выискивать себе домики, где в данный момент нет других самцов). Для самок RCS вычислялся аналогичным образом по «женским» RFID-сигналам. Поскольку самки, по-видимому, не особо конкурировали друг с другом в ходе эксперимента, «женский» RCS вряд ли отражает их конкурентоспособность. Однако для обоих полов RCS, вероятно, отражает доступ к пище.
Анализ динамики RCS подтвердил предположения авторов. Оказалось, что у самцов различия по RCS, во-первых, больше, чем у самок, во-вторых, растут с возрастом, в то время как у самок эти различия с возрастом не увеличиваются. Иными словами, среди самцов постепенно выделяются доминантные особи, имеющие эксклюзивный доступ к ресурсам, а у самок ничего подобного не происходит. Кроме того, выяснилось, что у самцов начиная примерно с 25–30-дневного возраста RCS начинает положительно коррелировать с массой тела. Иными словами, самцы, которым повезло занять лидирущие позиции в социуме, быстрее набирали вес. Разница в весе между лидерами и отстающими продолжала увеличиваться с возрастом. У самок этого не наблюдалось: самки с высокими значениями RCS в течение всего эксперимента не превосходили по массе тела самок с низкими значениями данного показателя.
Таким образом, результаты согласуются с идеей о том, что случайные различия, возникающие в начале жизни благодаря слепому везению, в дальнейшем могут усиливаться под влиянием конкуренции, выводя индивидов на самоподдерживающиеся, расходящиеся траектории развития.
Авторы отмечают, что в других экспериментальных условиях самки мышей тоже могли бы конкурировать друг с другом, и тогда у них, наверное, тоже наблюдалось бы «автокаталитическое» нарастание различий с возрастом.
Исследование подкрепляет модную ныне идею о том, что жизненный успех индивидов может во многом объясняться не их личными качествами, талантами и заслугами, а слепым везением. Это несколько подрывает устоявшиеся идеи о преимуществах меритократии (в социальных науках) и о всесилии полового отбора (в эволюционной биологии). Ведь если фенотипическая изменчивость в значительной степени определяется случайностью, а не генетическими различиями между индивидами, эффективность отбора будет ниже. Авторы даже предполагают, что это может быть одним из объяснений пресловутого «парадокса токовища» (lek paradox), который заключается в том, что даже сильный половой отбор не может убрать из генофонда всю изменчивость по генам, влияющим на сексуально привлекательные признаки, и сделать всех индивидов одинаково неотразимыми (см. Привередливость самок дрозофил помогает очищать генофонд от мутационного груза, «Элементы», 29.05.2018).
В больших популяциях на длительных временных интервалах эффекты везения должны усредняться и нивелировать друг друга, а гены, повышающие дарвиновскую приспособленность (включая успешность в конкуренции за половых партнеров), все равно будут закрепляться под действием отбора. Однако после того, как они закрепятся, вся оставшаяся фенотипическая изменчивость по признакам, влияющим на приспособленность, будет зависеть от удачи, а не от генов. В этом случае отбор окончательно утратит эффективность.
В заключение замечу, что статья оставляет двойственное впечатление. С одной стороны, результаты интересны и заставляют задуматься о важных теоретических вопросах. С другой, экспериментальные работы, публикуемые в журналах уровня Science, обычно выглядят весомее. В данном случае мы имеем единичный (без повторностей) эксперимент, проводившийся на сотне мышей в течение двух месяцев. Собранные данные (RFID-идентификации) позволяют оценивать поведенческие признаки и конкурентоспособность индивидов лишь косвенно, при помощи изощренной статистики. Мышам не дали даже произвести собственное потомство, что позволило бы посмотреть, каким образом выявленные поведенческие особенности транслируются в репродуктивный успех. Есть и мелкие неряшливости вроде приведенной в дополнительных материалах фотографии экспериментальной площадки, которая на самом деле показывает не ту площадку, которая использовалась в эксперименте, а какую-то другую (рис. 1). Имеющиеся в статье рассуждения о социальных науках и «эффекте Матфея» подразумевают, что у людей всё может быть так же, как у мышей в эксперименте, хотя основания для такого допущения в статье не обсуждаются. В сочетании с модностью и даже, осмелюсь сказать, идеологической нагруженностью главной идеи («неравенство идет не от талантов и заслуг, а от слепой удачи»), всё это, на мой взгляд, несколько снижает убедительность выводов исследования.
Источник: Matthew N. Zipple, Daniel Chang Kuo, Xinmiao Meng, Tess M. Reichard, Kwynn Guess, Caleb C. Vogt, Andrew H. Moeller, Michael J. Sheehan. Competitive social feedback amplifies the role of early life contingency in male mice // Science. 2025. DOI: 10.1126/science.adq0579.
Свежие комментарии