

Латимерия — один из немногих современных представителей группы мясистолопастных рыб, среди которой были предки наземных позвоночных. Кроме нее, к мясистолопастным относятся двоякодышащие рыбы, у которых, как и у наземных позвоночных, есть настоящие легкие. А что есть у латимерии — легкие, плавательный пузырь или ни то, ни другое? Как ни удивительно, на протяжении нескольких десятилетий на этот вопрос не было ясного ответа. Судя по недавно полученным данным, у латимерии есть маленькое рудиментарное (утратившее дыхательную функцию) непарное легкое, которое снабжается кровью через легочные артерии — так же, как и у других обладателей легких.
Чем отличаются рыбы от наземных позвоночных? Одно из очевидных отличий, которые назовет кто угодно, заключается в способе дыхания: рыбы дышат жабрами, а наземные позвоночные — легкими. Когда в середине XIX века были открыты двоякодышащие рыбы, имеющие настоящие легкие, зоологи сначала приняли их за гигантских амфибий. Правда, при ближайшем рассмотрении оказалось, что многие другие признаки этих животных заставляют отнести их все-таки к рыбам. Но дискуссия по этому вопросу была напряженной и заняла не одно десятилетие. Ее памятником служит латинское название южноамериканской двоякодышащей рыбы — Lepidosiren paradoxa, что можно перевести как «парадоксальный чешуйчатый сирен» (сирены — это водные хвостатые амфибии, на которых лепидосирен внешне отдаленно похож). Другие современные двоякодышащие рыбы — это африканский протоптер (Protopterus, рис. 2) и австралийский рогозуб (Neoceratodus).
Кроме них, легкие есть у многоперообразных рыб, живущих в Африке и представленных родами Polypterus и Erpetoichthys, и у наземных позвоночных. Последние, таким образом, вовсе не уникальны по этому признаку.
Появление легких не было связано с выходом на сушу. И двоякодышащие, и многоперообразные пользуются воздушным дыханием, живя в воде. Более того, лепидосирен и протоптер живут в водоемах, которые время от времени пересыхают. Но даже тогда они не выползают на сушу, а зарываются в дно водоема и впадают в летнюю спячку. Рогозуб, который живет в непересыхающих реках, не делает и этого.
Как и зачем возникли легкие? Это весьма непростые вопросы, которые стоит рассмотреть по очереди.
Легкие и плавательный пузырь: гомологи
Легкие — это парные дыхательные мешки, которые образуются в качестве выростов нижней (наиболее близкой к брюху) части пищевода. В свою очередь, пищевод является продолжением глотки — передней части кишечной трубки, которая у водных позвоночных образует ряд парных жаберных мешков, открывающихся наружу жаберными щелями. Раньше считалось, что дыхательный эпителий жабр у бесчелюстных позвоночных образован внутренним зародышевым листком (энтодермой), а у рыб внешним зародышевым листком (эктодермой). Но исследования экспрессии генов развития показали, что жабры и у бесчелюстных, и у рыб имеют одинаковое происхождение — энтодермальное (J. Gillis, O. Tidswell, 2017. The origin of vertebrate gills). Другими словами, они происходят в конечном счете из кишечной трубки. Легкие — тоже энтодермальные органы, происходящие из кишечной трубки. Еще в 1875 году немецкий биолог Александр Гетте (Alexander Wilhelm von Goette) предположил, исходя из эмбриологических данных, что легкие развились из самой задней пары жаберных мешков, которые модифицировались и не стали прорываться наружу. Действительно, у современных амфибий, причем у всех трех групп — бесхвостых, хвостатых и безногих, — легкие начинают развиваться так, как если бы это были зачатки жаберных мешков (S. Perry et al., 2001. Which came first, the lung or the breath?). Допустить, что это отражает их эволюционное происхождение, очень легко.
К проблеме происхождения легких примыкает проблема происхождения плавательного пузыря. В отличие от парных легких, являющихся выростами брюшной части пищевода, плавательный пузырь всегда непарный и является выростом спинной части пищевода (рис. 3). Он может иметь дыхательную функцию, но может ее и не иметь. К обладателям респираторного плавательного пузыря относятся, например, панцирная щука (см. Архаичные гены костных ганоидов разнообразнее, чем у более молодых групп позвоночных, «Элементы», 11.07.2016) и амия (см. картинку дня Амия, рыба из юрского периода). Но в большинстве случаев основной становится гидродинамическая функция плавательного пузыря: газ, которым он заполнен, легче воды, и это помогает рыбе не тонуть. Как заметили британские палеонтологи Майкл Пирсон (Michael Pearson) и Томас Вестолл (Thomas Stanley Westoll), «вопрос о его [плавательного пузыря] исходной функции, дыхательной или гидродинамической, лишен смысла, поскольку любое газовое тело в глотке немедленно оказывает влияние на плавучесть» (M. Pearson, T. S. Westoll, 1979. The Devonian actinopterygian Cheirolepis Agassiz). У многих рыб — например, у окунеобразных — сформировалось так называемое закрытопузырное состояние, когда проток, связывающий плавательный пузырь с пищеводом (ductus pneumaticus), у взрослой рыбы полностью закрывается. В этом случае дыхательная функция, разумеется, исключена.
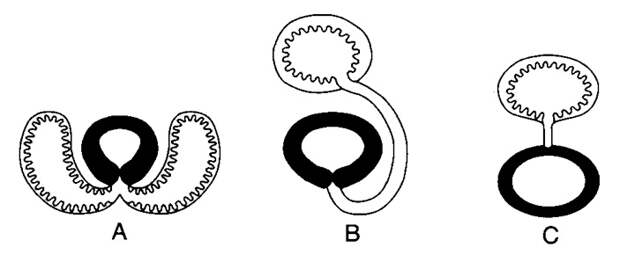
У кого есть плавательный пузырь? Чтобы корректно ответить на этот вопрос, нам нужно вспомнить, что все костные рыбы делятся на два эволюционных ствола — мясистолопастные (они же лопастеперые) и лучеперые. Двоякодышащие, например, относятся к мясистолопастным. А вот многоперообразные относятся к лучеперым, среди которых они образуют самую древнюю (базальную) эволюционную ветвь. Группа, в которую входят все лучеперые, за исключением многоперообразных, называется Actinopteri. Вот эта группа и объединяет всех обладателей плавательного пузыря. Ни у кого, кроме Actinopteri, такой орган не встречается. Зато среди Actinopteri он есть у всех, если не считать случаев вторичной редукции. Плавательный пузырь может исчезать или у донных рыб вроде камбалы, или у постоянно быстро плавающих вроде тунца; но нет сомнений, что у предков этих рыб он был. Есть все основания считать, что плавательный пузырь возник в эволюции только один раз — у общего предка группы Actinopteri.
Какое отношение имеет плавательный пузырь к легким? Вопреки иногда встречающимся утверждениям, сейчас практически никто не считает, что легкие произошли от плавательного пузыря. Конфигурация эволюционного древа указывает на обратное (рис. 4). Легкие — это очень древние органы. Помимо костных рыб, к которым относятся двоякодышащие и многоперообразные, остатки легких были описаны у древней панцирной рыбы Bothriolepis (R. Denison, 1941. The soft anatomy of Bothriolepis). Иногда это открытие, как водится, ставят под сомнение (D. Goujet, 2011. „Lungs“ in Placoderms, a persistent palaeobiological myth related to environmental preconceived interpretations), но вероятность, что речь идет о легких, все же не следует сбрасывать со счета. В любом случае решить вопрос об отношениях легких и плавательного пузыря можно и без ботриолеписа. Присутствие легких у двоякодышащих и многоперообразных — двух далеких друг от друга ветвей, одна из которых располагается среди мясистолопастных, а другая в основании ствола лучеперых — однозначно указывает на то, что легкие возникли раньше плавательного пузыря. Еще в 1895 году американский биолог Башфорд Дин (Bashford Dean) опубликовал гипотезу, что плавательный пузырь произошел от легкого, которое стало непарным и сместилось с брюшной стороны на спинную. Это выглядит логично: гидродинамический орган, содержащий газ, который легче воды, лучше расположить выше центра тяжести.
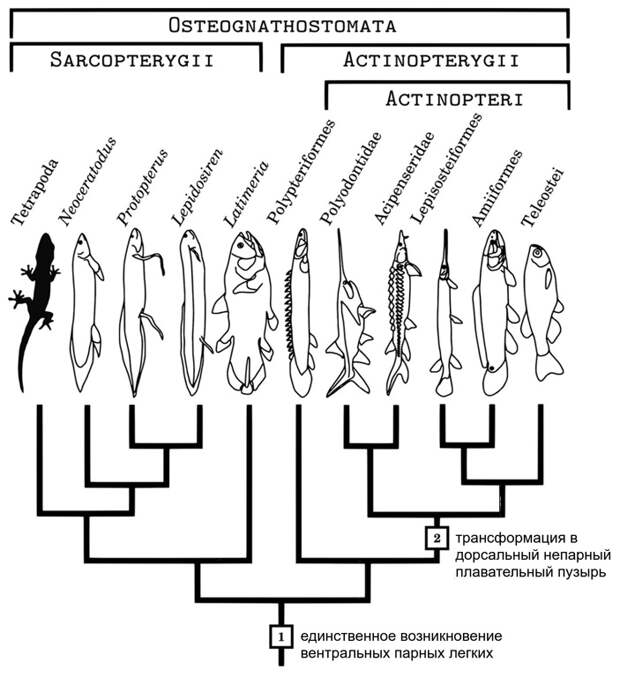
Современная генетика развития подтверждает родство легких и плавательного пузыря. Описана генная сеть из шести регуляторных генов, которая управляет эмбриональным развитием как плавательного пузыря данио-рерио, так и легких мыши (A. Cass et al., 2013. Expression of a lung developmental cassette in the adult and developing zebrafish swimbladder). Исследования транскриптомов тоже показывают, что по набору генов, задействованных в их развитии, плавательный пузырь рыб и легкие млекопитающих близки друг к другу (W. Zheng et al., 2011. Comparative transcriptome analyses indicate molecular homology of zebrafish swimbladder and mammalian lung). Правда, о том, в каком порядке эти органы возникали в эволюции, генетика развития ничего сказать не может.
Тем не менее имеющихся фактов вполне хватает, чтобы объединить гипотезы Гетте и Дина, признав их в целом верными. Некогда у древнейших костных рыб самая задняя пара жаберных мешков перестала прорываться наружу и превратилась в слепо замкнутые органы воздушного дыхания — легкие. В дальнейшем, когда обстановка в водоемах изменилась, потребность в органах воздушного дыхания уменьшилась. Легкие утратили парность и переместились из подкишечного положения в надкишечное, чтобы служить гидродинамическим органом — плавательным пузырем. Так выглядит широко распространенный сценарий, который вошел во многие современные учебники (например, в книгу Кеннета Кардонга «Vertebrates: comparative anatomy, function, evolution»).
...или нет?
Однако у этого сценария есть и проблемы. Между плавательным пузырем и легкими нет реальных переходных этапов. Попытки найти их оказались несостоятельными (K. Liem, 1988. Form and function of lungs: the evolution of air breathing mechanisms). Отверстие, ведущее в легкие / плавательный пузырь, находится или с брюшной стороны пищевода, или со спинной. Надо заметить, что у австралийской двоякодышащей рыбы — рогозуба — легкое таки действительно сместилось на спинную сторону и стало непарным (см. рис. 3). Но ведущий в это легкое проток огибает пищевод справа и впадает в него по-прежнему с брюшной стороны. Миграции отверстия воздушного протока на спинную сторону у рогозуба не произошло, и нет подтверждений тому, что такая миграция происходила в ходе эволюции у кого бы то ни было.
В принципе можно было бы допустить, что у каких-то рыб произошел поворот отрезка кишечной трубки на 180 градусов, так что орган воздушного дыхания, впадавший в пищевод снизу, оказался сверху. Но подобные повороты обычно оставляют следы в прохождении сосудов и нервов, чего в нашем случае нет. Поэтому гипотезу о повороте кишечной трубки никто всерьез не рассматривает.
Посмотрим на проблему с теоретической точки зрения. В случае, когда один орган в ходе эволюции превратился в другой орган, мы говорим, что эти органы гомологичны. Существует три основных критерия гомологии: критерий положения, критерий непрерывности и критерий специальных качеств. Гомология легких и плавательного пузыря не подтверждается ни критерием положения (они расположены по-разному), ни критерием непрерывности (между ними нет переходных состояний). Значит, полагаться можно только на критерий специальных качеств. Действительно, плавательный пузырь и легкие близки друг другу как по структуре газообменных тканей, так и по экспрессии генов развития. Вопрос в том, насколько эти качества уникальны. По современным рыбам мы знаем, что органы воздушного дыхания могут возникать в ходе эволюции независимо и многократно. Таких примеров масса: лабиринтовые, некоторые сомы, змееголовы, обыкновенный вьюн и так далее. Естественно, что в органах, выполняющих одну функцию, реализуются примерно одни и те же «технологические решения». Такие вещи широко распространены в живой природе. Есть данные, что одни и те же регуляторные гены и сигнальные белки задействованы даже в развитии легких мыши и трахей мухи-дрозофилы (M. Behr, 2010. Molecular aspects of respiratory and vascular tube development). Легкие млекопитающих и трахеи насекомых — это органы дыхания, для структурных элементов которых характерно ветвление (отсюда и общие гены), но по происхождению они не имеют между собой ничего общего. Решающим аргументом для установления гомологии экспрессия общих генов в таких случаях служить не может.
Альтернативная идея была предложена в большой серии статей, одним из авторов которых неизменно выступал американский биолог Стивен Перри (Steven Franklin Perry). Еще в 1932 году русский биолог Владимир Викторович Васнецов показал, что у зародыша стерляди позади ряда жаберных мешков глотка образует серию дополнительных выростов, которые выглядят как заготовки для будущих органов дыхания (рис. 5). Этих выростов пять: один непарный спинной (дорсальный) и две пары боковых (дорсолатеральные и вентролатеральные). Стерлядь — обладатель типичного плавательного пузыря (функцию дыхания он у осетрообразных уже утратил). Этот плавательный пузырь образуется из непарного спинного выроста глотки. Парные боковые выросты, из которых могли бы образоваться легкие, никаких органов взрослого животного не дают.
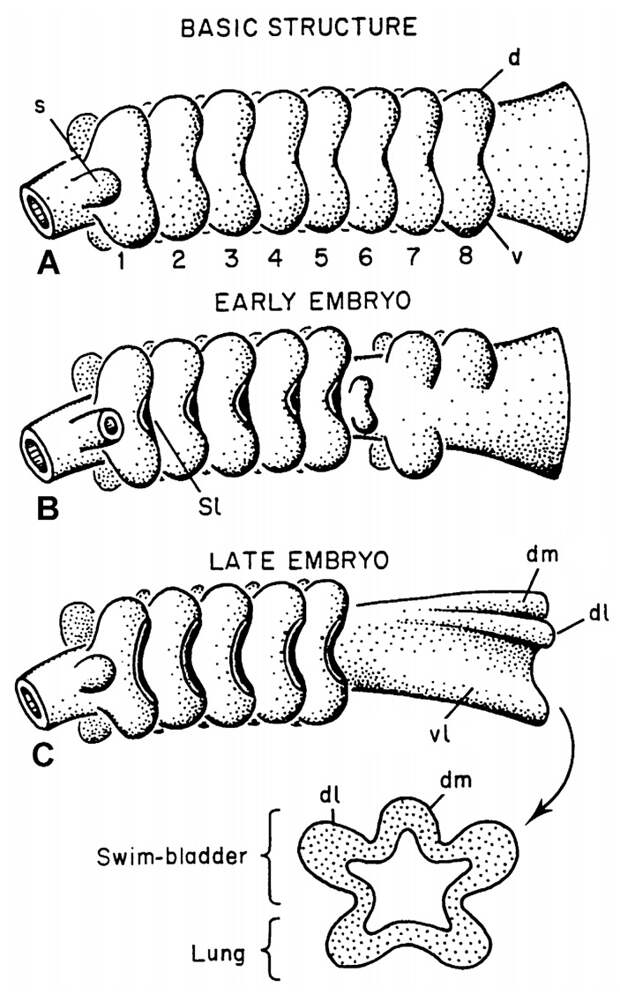
Наблюдения Васнецова противоречат гомологии плавательного пузыря и легких, зато делают возможным другой эволюционный сценарий. По мнению Перри, у древнейших костных рыб позади ряда жаберных мешков находился отдел глотки, предназначенный для воздушного дыхания — респираторная глотка (respiratory pharynx). Для увеличения дыхательной поверхности респираторная глотка имела пять выростов — непарный спинной и две пары боковых, так что в поперечном сечении она выглядела бы пятилучевой. В разных группах рыб судьба разных частей респираторной глотки была различна. У двоякодышащих и у многоперообразных сильно развились нижние боковые (вентролатеральные) выросты глотки, которые по положению точно соответствуют легким; очевидно, что они и были предшественниками легких. Как мы уже знаем, у стерляди из этих выростов ничего не развивается. Зато в группе Actinopteri (стерлядь относится именно к ней) сильно развился непарный спинной вырост глотки, который и дал плавательный пузырь. Таким образом, легкие и плавательный пузырь возникли из недифференцированной респираторной глотки независимо (рис. 6).
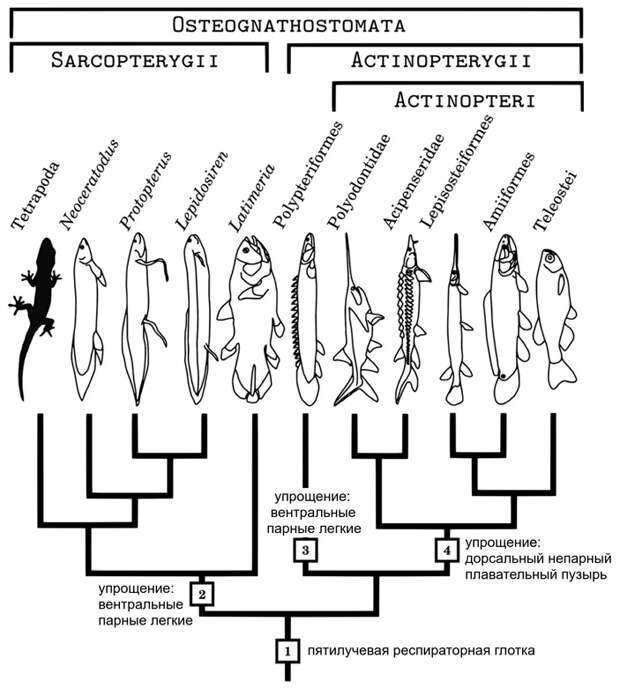
К этому можно добавить указание Бронислава Александровича Домбровского, что легкие и плавательный пузырь находятся по разные стороны перикардно-перитонеальной перегородки (Б. А. Домбровский, 1982. Сравнительная морфология животных и синтетическая зоология). Эта перегородка, сходная с диафрагмой млекопитающих, отделяет у рыб околосердечную полость от брюшной. Легкие находятся впереди от нее, а плавательный пузырь — позади. Очевидно, это должно означать, что плавательный пузырь развился не из того же сегмента глотки, что и легкие, а из более заднего. Это опять же противоречит гомологии плавательного пузыря и легких, но не противоречит гипотезе Перри. Правда, гипотеза Перри выглядит сложнее классической гипотезы Дина, но это слабый довод: живая природа вовсе не обязана быть простой.
Загадка второго круга
Важную часть проблемы происхождения легких составляет проблема их кровоснабжения. Все мы знаем, что одной из главных функций кровеносной системы является перенос кислорода. Для начала представим себе «нормальную» рыбу, получающую кислород в основном из жабр (рис. 7). В сердце такой рыбы поступает венозная кровь, уже отдавшая кислород многочисленным органам и тканям тела. Сокращение желудочка сердца выбрасывает эту кровь в брюшную аорту, которая проходит прямо под глоткой. От брюшной аорты отходят приносящие жаберные артерии, которые несут венозную кровь в жабры. В жабрах каждая приносящая жаберная артерия распадается на многочисленные тончайшие капилляры, в которые поступает кислород из омывающей жаберные лепестки воды. Там венозная кровь насыщается кислородом, то есть, другими словами, становится артериальной. Из жаберных капилляров артериальная кровь собирается в выносящие жаберные артерии, которые, в свою очередь, впадают в корни спинной аорты — парные крупные сосуды, проходящие над глоткой. Сзади корни спинной аорты сливаются в непарную спинную аорту, которая несет артериальную кровь к туловищу, а впереди продолжаются в сонные артерии, которые несут артериальную кровь в голову. Приносящая и выносящая жаберные артерии, обслуживающие одну и ту же жабру, называются артериальной дугой (запомним этот термин, он нам пригодится).
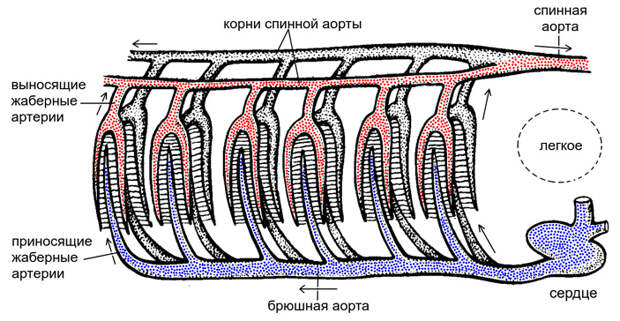
Обратим внимание: в кровеносной системе, которую мы только что описали, нет никакого смешивания артериальной и венозной крови. Такое смешивание появляется, когда вместе с легкими возникает второй круг кровообращения: в этом случае артериальная кровь от легких поступает прямо в сердце, «разбавляя» венозную кровь, поступающую от всего остального тела. Смешивание артериальной и венозной крови создает множество физиологических проблем. С одной стороны, жабры вместо чисто венозной крови теперь будут получать смешанную, уже содержащую какое-то количество кислорода из легких. В результате снизится эффективность газообмена в жабрах — ведь она тем выше, чем больше разность концентраций кислорода в крови и в омывающей жабры воде. Если в воде кислорода мало, эта разность может вообще поменять знак, и тогда жабры будут отдавать кислород из крови в воду, а не поглощать его (К. Шмидт-Ниельсен, 1982. Физиология животных: приспособление и среда). Все остальное тело, в свою очередь, будет получать вместо чисто артериальной крови в той или иной мере смешанную, что, естественно, снизит ее энергетическую ценность. Конечно, эти проблемы можно свести к минимуму, вводя в действие всевозможные хитрые механизмы вторичного разделения артериальной и венозной крови. Но разделить их полностью удалось (из современных животных) только птицам и млекопитающим.
А нельзя ли было подключить легкое к кровеносной системе таким образом, чтобы никакого смешивания крови не возникло? Легкие, как мы знаем, происходят от задней недоразвитой пары жаберных мешков. Это означает, что легкое находится прямо позади ряда нормальных жабр. Решение, которое на первый взгляд может показаться простым и эффективным, состоит в том, чтобы подключить легкое к кровеносной системе параллельно — по примеру жабр, через дополнительную приносящую и дополнительную выносящую жаберные артерии. С эмбриологической точки зрения это легко сделать, и никакого смешивания артериальной крови с венозной в этом случае не будет. Круг кровообращения останется единственным. Однако физиологически такая система будет неработоспособной. Почему?
Ответ на этот вопрос дал в 1960 году Леонид Петрович Татаринов. Дело в том, что в рыбьей кровеносной системе, изначально «настроенной» на серию жабр, разные артериальные дуги имеют разное функциональное значение. Мы ведь помним, что артериальная кровь, получившая кислород в жаберном аппарате, должна идти как вперед — через сонные артерии в голову, так и назад — через спинную аорту в туловище. Благодаря физиологическим исследованиям известно, что из двух самых передних артериальных дуг кровь идет в голову, а из двух самых задних — в туловище. Теперь представим, что мы подключили легкое к кровеносной системе через дополнительную артериальную дугу. Кровь из нее пойдет в спинную аорту и дальше в туловище — пространственное расположение не позволяет иного. Если такое животное полностью перейдет на легочное дыхание, то его голова не получит ни капли артериальной крови. А ведь из той же физиологии хорошо известно, что нет органа, более требовательного по части кислорода, чем головной мозг. Если орган дыхания бесполезен для снабжения мозга кислородом — значит, он бесполезен вообще.
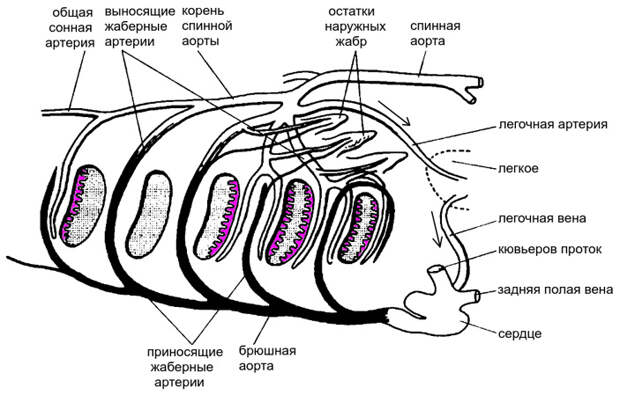
Вот почему легкое пришлось подключить к кровеносной системе не параллельно, а последовательно. У реальных обладателей легких легочная артерия отходит не от приносящей, а от выносящей части последней артериальной дуги (рис. 8). Это означает, что легкое получает кровь, уже прошедшую через жабры, чтобы донасытить ее кислородом. Чем хуже работают жабры, тем важнее становится роль легкого. А из легкого кровь поступает по легочной вене прямо в сердце. Это единственный способ донести кислород из легкого до всего тела. Выигрыш от наличия легкого, очевидно, оказался выше, чем проигрыш от побочных эффектов, вызванных появлением второго круга.
Гипотеза Фармер
Несколько иную интерпретацию эволюционной роли легких предложила американская исследовательница Колин Фармер (Colleen Farmer). Она начала с вопроса: зачем, собственно, легкие понадобились древним рыбам? Зададимся этим вопросом и мы.
Версию, что легкие появились для экскурсий на сушу, пришлось отбросить сразу же, как только выяснилось, что современные двоякодышащие рыбы не проявляют никакого желания даже на время покидать воду. Стало ясно, что, как бы древние рыбы ни использовали легкие, они делали это в воде. Здесь на помощь пришла палеонтология. Есть все основания считать, что для девонских пресных водоемов (не говоря уж о более ранних) было характерно низкое содержание кислорода в воде, а главное — его резкие колебания, поскольку экологическая обстановка в этих водоемах была очень нестабильной (см. картинку дня Ризодус). В то же время эти водоемы были мелкими: рыбам хватало небольшого мускульного усилия, чтобы подняться к поверхности и глотнуть воздуха. Налицо была как потребность в органе воздушного дыхания, так и удобная возможность его использовать. Итак, легкие возникли у древних рыб в качестве приспособления к гипоксии. Другим вариантом такого приспособления могло бы быть простое увеличение числа жаберных мешков, как у бесчелюстных из группы галеаспид. Но в атмосферном воздухе кислорода больше, чем в воде, и в качестве «запасного» органа дыхания легкое эффективнее.
Фармер, однако, обратила внимание на то, что данные о реальных обладателях легких не вполне подтверждают эти выводы. Например, современный австралийский рогозуб живет в довольно глубоких реках, где никакой существенной гипоксии не наблюдается даже в летнюю межень, при наибольшем снижении уровня воды (G. Grigg, 1965. Studies on the Queensland lungfish, Neoceratodus forsteri (Krefft): III. Aerial respiration in relation to habits). Наблюдения, сопровожденные лабораторными экспериментами, показали, что «триггером», запускающим у рогозуба воздушное дыхание, служит не внешняя обстановка, а двигательная активность самой рыбы. Когда рогозуб охотится (что в природе обычно бывает ночью), у него за счет работы мышц подскакивает потребность в кислороде, и он удовлетворяет ее, поднимаясь к поверхности и глотая воздух. Но этот способ дыхания всегда остается дополнительным; полностью перейти на легочное дыхание рогозуб неспособен, и в летнюю спячку он, в отличие от протоптера и лепидосирена, никогда не впадает. Тут надо заметить, что рогозуб — самая примитивная двоякодышащая рыба из доживших до наших дней. По морфологическим признакам, например по строению плавников и чешуй, он гораздо ближе к девонским предкам, чем протоптер и лепидосирен. Что, если и с типом дыхания дело обстоит так же?
Очень похожие данные есть и для многопера (Polypterus), исключительно примитивной лучеперой рыбы, обладающей типичными легкими. Эксперименты показали, что частота воздушного дыхания у многопера зависит от мышечной активности гораздо сильнее, чем от содержания кислорода в воде: «Активная рыба в аэрированной воде поднималась к поверхности чаще, чем неактивная рыба в неаэрированной воде» (A. Magid, 1966. Breathing and function of the spiracles in Polypterus senegalus).
Итак, не стоит ли поискать какую-то другую причину возникновения легких, кроме внешних гипоксических условий?
Фармер считает, что такой причиной могло послужить снабжение кислородом сердечной мышцы. Мышечный слой стенки сердца, как известно, называется миокардом. Для водных позвоночных — бесчелюстных и рыб — характерен губчатый миокард, где между тяжами мышечных клеток есть многочисленные свободные пространства. Это позволяет снабжать миокард кислородом прямо из крови, заполняющей сердце. Мы помним, что у обладателей одного круга кровообращения через сердце проходит только венозная кровь. Но даже венозная кровь все же содержит какое-то количество кислорода, причем его там тем больше, чем лучше у данного животного развито кожное дыхание. У личинки миноги — пескоройки — кожное дыхание при ее маленьком размере развито очень хорошо, и кровоснабжение сердца не вызывает никаких проблем (рис. 9, a). Взрослой миноге, которая дышит уже в основном жабрами, приходится сложнее, но и она с этой задачей справляется.
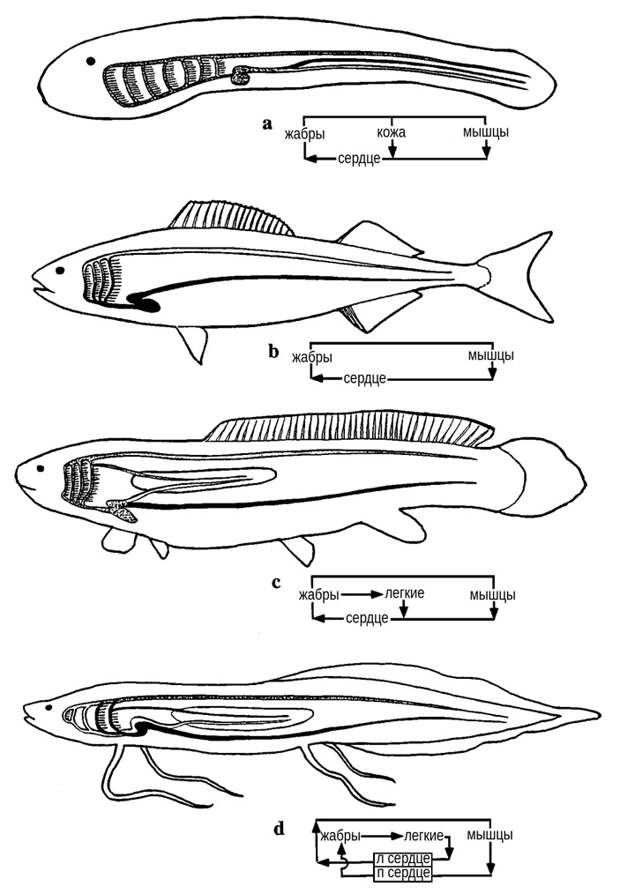
Более продвинутый способ кровоснабжения миокарда заключается в том, чтобы пронизать его специальными тонкими артериями. Эти артерии называются коронарными. Но они появились далеко не сразу. У бесчелюстных — миног и миксин — никаких коронарных сосудов просто нет. У этих существ сердце может снабжаться кислородом только из крови, заполняющей его собственный просвет. У рыб система коронарных сосудов начала формироваться, но этот процесс был непростым и постепенным. Между тем рыбы — гораздо более подвижные существа, чем бесчелюстные, и потребность в кислороде у них намного выше. И вот тут на помощь пришло легкое. По легочной вене артериальная кровь, максимально насыщенная кислородом, впрыскивается прямо в сердце, обеспечивая оксигенацию миокарда (рис. 9, b-c).
Соображения Фармер подтверждаются некоторыми физиологическими данными. Например, у амии (которая, как мы помним, имеет респираторный плавательный пузырь — полный функциональный аналог легких) начинаются нарушения работы сердца, если заставить ее активно плавать, лишив доступа к атмосферному воздуху; если доступ к воздуху есть, та же нагрузка не вызывает никаких проблем.
С этой точки зрения смешивание артериальной и венозной крови начинает выглядеть не побочным эффектом, а вполне себе адаптацией. Становится ясно, почему у большинства обладателей легких и респираторных плавательных пузырей — у амии, панцирной щуки, многопера, рогозуба — перегородки между артериальной и венозной частью сердца либо отсутствуют, либо развиты очень слабо. Смешивание крови — «не баг, а фича», оно поддерживается, пока польза от него превышает вред.
Особое положение на этом фоне занимают африканские и южноамериканские двоякодышащие рыбы — протоптер и лепидосирен (рис. 9, d). Вот у них потоки артериальной и венозной крови разделены (в предсердии есть служащая для этого перегородка). Кроме того, у них частично атрофированы жабры, чтобы предотвратить потерю кислорода, если в окружающей воде его окажется меньше, чем в проходящей сквозь жабры крови (см. рис. 8). Эти рыбы действительно хорошо адаптированы к гипоксическим условиям. Но была ли такая адаптация изначально свойственна всем обладателям легких? Фармер уверена, что нет. Протоптер и лепидосирен — сильно специализированные формы. Их предки, скорее всего, были по характеру дыхания ближе к более примитивным рыбам — рогозубу и многоперу.
Может быть, Фармер и преувеличивает, называя оксигенацию миокарда единственной причиной появления легких. Этого мы пока не знаем. Но ее работы помогают нам понять, что биологическая адаптация всегда многогранна, и увидеть грань, которую мы раньше не видели.
Легкое латимерии
В мозаике, которую мы сложили, не хватает одного важного фрагмента. Кроме двоякодышащих, к группе мясистолопастных относится род латимерия (Latimeria; рис. 1). Этот род представлен двумя современными видами: коморская латимерия (Latimeria chalumnae) и индонезийская латимерия (Latimeria menadoensis). Первый вид живет на западе Индийского океана, а второй — в море Сулавеси, которое относится уже к Тихому океану.
Открытие латимерии, как известно, произошло в 1938 году, когда близ Ист-Лондона, на самом юго-западе Индийского океана, рыбаки выловили странную рыбу. Сообщение об этой рыбе получил южноафриканский ихтиолог Джеймс Леонард Брайерли Смит (James Leonard Brierley Smith). Осмотрев ее, Смит сразу сделал абсолютно правильный вывод: рыбу следует отнести к отряду Actinistia, к семейству Coelacanthidae, которое считалось вымершим в мезозое (J. L. B. Smith, 1939. A living fish of mesozoic type). Целаканты, они же актинистии, были хорошо известны палеонтологам еще в XIX веке. Семейство Coelacanthidae выделил в 1843 году Жан Луи Агассис (Jean Louis Rodolphe Agassiz), а отряд Actinistia выделил в 1871 году Эдвард Коп (Edward Drinker Cope) (кстати, в той же работе Коп ввел и упомянутый выше таксон Actinopteri; E. D. Cope, 1871. Contribution to the ichthyology of the Lesser Antilles). Но никто не знал, что эти древние рыбы дожили до наших дней.
Обсуждая латимерию, премьер-министр Южно-Африканского Союза Даниэль Франсуа Малан (Daniël François Malan) спросил Смита: «Вы хотите сказать, что и мы некогда выглядели таким образом?» Смит несколько смутился и отделался шуткой: «Гм... Я встречал людей и пострашнее». Известный английский биолог Джулиан Хаксли (Julian Sorell Huxley) потом сказал, что на самом деле следовало бы ответить: «Вообще говоря, да!»
Верно ли это? С одной стороны, целаканты не являются предками человека. Это особая ветвь мясистолопастных рыб, гораздо более далекая от наземных позвоночных, чем двоякодышащие рыбы (рогозуб, протоптер и лепидосирен). Этот вывод, уже сделанный палеонтологами (D. Rosen et al., 1981. Lungfishes, tetrapods, paleontology, and plesiomorphy), в начале XXI века подтвердила молекулярная филогенетика (D. Liang et al., 2013. One thousand two hundred ninety nuclear genes from a genome-wide survey support lungfishes as the sister group of tetrapods). В строении черепа, плавников, хвоста целакантов хватает уникальных признаков, которых у предков наземных позвоночных никогда не было. С другой стороны, целаканты — это все же настоящие мясистолопастные рыбы, сохранившие древний облик и многие архаичные черты. Поэтому латимерия может дать чрезвычайно важные подсказки, касающиеся хода эволюции — в том числе и на пути к человеку.
Латимерия живет в море на глубинах около 200 метров, иногда опускаясь даже до 700 метров. Ни о каком воздушном дыхании в этих условиях не может быть и речи. Тем не менее у латимерии есть рудиментарное легкое. Оно представляет собой вытянутый мешок, расположенный под пищеводом и соединенный с ним протоком, который остается открытым на протяжении всей жизни рыбы. Этот орган очень мал: у 130-сантиметровой рыбины его длина, если не считать суженной в ниточку задней части, составляет меньше трех с половиной сантиметров (рис. 10). Перед нами именно рудимент — остаток органа, который давно потерял свою первоначальную функцию. Внутренняя поверхность легкого латимерии имеет складки, которые когда-то, вероятно, служили для увеличения дыхательной поверхности. А его внешняя соединительно-тканная оболочка содержит плотные слои жира; к тому же легкое погружено в расположенную под кишечной трубкой латимерии массу жира, которая называется жировым органом. Раньше этот жировой орган часто принимали за плавательный пузырь или «жировое легкое». Латимерия вообще необычайно склонна к накоплению в своих тканях жира или масла (Смит, например, писал, что на поверхности консервирующей жидкости, в которой хранился один из экземпляров латимерии, постоянно плавали большие шарики жира). Очевидно, это способствует плавучести — ведь жир и масло легче воды. Например, у акул, у которых, как и у всех хрящевых рыб, нет плавательного пузыря, такую же роль играет огромная богатая липидами печень (C. Phleger, 1998. Buoyancy in marine fishes: direct and indirect role of lipids).

Гораздо более крупное легкое известно у ископаемых целакантов. Так же, как и легкое латимерии, оно всегда непарное. Легкое целакантов — редкий случай — хорошо отображается на каменных отпечатках, потому что оно со всех сторон окружено костными пластинками (рис. 11). У латимерии тоже есть похожие пластинки, но у нее они, как и само легкое, рудиментарны.

Парное или непарное?
Как можно интерпретировать этот орган? Учтем, что оба вида латимерии малочисленны, уязвимы и находятся под охраной. Поэтому возможность вскрыть латимерию выпадает биологам очень редко. Кроме того, латимерия — рыба живородящая, а это крайне затрудняет любые эмбриологические исследования. Жаловаться тут не на что: сохранить удивительный редкий вид животных, конечно, важнее, чем получить научную информацию. Сейчас можно лишь констатировать: насколько нам известно, легкое целакантов не проявляет никаких следов парности.
У двусторонне-симметричных животных, в том числе и у позвоночных, истинно непарные органы встречаются редко. Зато бывает, что парный орган постепенно переходит в непарное состояние. Именно это, судя по всему, произошло с легкими рогозуба: у него сохраняется только правое легкое, а левое появляется в ходе развития, но потом дегенерирует (M. Lambertz, 2017. The vestigial lung of the coelacanth and its implications for understanding pulmonary diversity among vertebrates: new perspectives and open questions). У большинства змей тоже сохраняется только правое легкое, что, разумеется, вызвано формой их тела. Процесс редукции одного из легких идет и у безногих ящериц, например у желтопузика (M. Lambertz et al., 2018. Variability in pulmonary reduction and asymmetry in a serpentiform lizard: the sheltopusik, Pseudopus apodus).
С другой стороны, плавательный пузырь — это, похоже, как раз пример истинно непарного органа. Гипотезу, что легкое целакантов тоже истинно непарное, пока нельзя как следует подтвердить, но нельзя и отбросить.
Бразильская исследовательница Камила Купелло (Camila Cupello) с коллегами обращают внимание на то, что признаки непарности легких есть не только у целакантов (C. Cupello et al., 2022. Lung evolution in vertebrates and the water-to-land transition). Например, у многопера легкие асимметричны: только правое легкое впадает в пищевод, а левое легкое впадает в правое. У эмбриона многопера легкие закладываются как непарный выступ на брюшной стороне глотки, который только потом разделяется на зачатки правого и левого легкого. Опираясь на эти и некоторые другие факты, Купелло и ее коллеги делают вывод, что легкое изначально было непарным органом, а его парность — производное состояние.
Однако принимать такую смелую гипотезу все же рановато. Данные, на которых она основана, допускают разные толкования. Ранняя эмбриональная закладка парных легких может быть непарной, но надо еще доказать, что такое состояние унаследовано от далеких предков, а не является вторичной модификацией хода развития. В конце концов, у амниот (рептилий, птиц и млекопитающих) ранняя закладка легких тоже непарная (S. Perry et al., 2001. Which came first, the lung or the breath?), но это никогда не было поводом считать, что у взрослых предков амниот легкое было непарным органом. А о начальных стадиях развития легкого латимерии пока ничего не известно. Версия, что его непарность вторична, все же не исключена.
Есть остроумное предположение, что второе легкое латимерии превратилось в жировой орган, разросшийся, но потерявший связь с пищеводом (M. Lambertz, 2017. The vestigial lung of the coelacanth and its implications for understanding pulmonary diversity among vertebrates: new perspectives and open questions). Правда, это противоречит тому, что у ископаемых целакантов легкое, которое, видимо, еще служило органом дыхания, тоже непарное.
А может быть, исходно непарное (вентромедиальное) легкое — это еще один, свойственный только целакантам, независимый способ выделения органа дыхания из недифференцированной респираторной глотки? Эта идея не противоречит ничему. К ней, судя по всему, склоняется немецкий биолог Маркус Ламбертц (Marcus Lambertz), друг и коллега ныне, к сожалению, покойного Стивена Перри. Если эта гипотеза верна, то парные легкие многоперов и двоякодышащих тоже должны были возникнуть независимо друг от друга. Но с точки зрения теории эволюции в этом нет ничего невероятного; более того, еще до появления новых данных о легком латимерии Ламбертц и Перри именно так и думали (см. рис. 6).
Легочные артерии
Редкость латимерии и сложность работы с ней порождают неожиданные пробелы в знаниях по ее анатомии и физиологии. Казалось бы, о таком знаменитом животном биологи должны знать все. Ничего подобного! Уже упоминалось, что рудиментарное легкое латимерии долгое время путали с жировым органом. А о кровоснабжении легкого латимерии до последнего времени было вообще ничего не известно. Мелькавшие в литературе описания ее легочных артерий оказались ошибкой: более тщательное исследование показало, что за сосуды были приняты затвердевшие элементы внешней оболочки легкого (C. Cupello et al., 2017. Lung anatomy and histology of the extant coelacanth shed light on the loss of air-breathing during deep-water adaptation in actinistians). Камила Купелло и ее коллеги очень тщательно, применяя метод рентгеновской компьютерной томографии, изучили пять экземпляров латимерии, которые хранятся в Музее естественной истории в Париже. В результате они получили много информации о строении легкого, но о его сосудах так и не смогли ничего сказать.
Недавно они заново исследовали один из «парижских» экземпляров латимерии — молодую самку длиной 42 сантиметра, пойманную в 1974 году у Коморских островов. Когда-то давно эту рыбу просвечивали рентгеном и ввели в ее кровеносную систему контрастное вещество — сульфат бария. Это ухудшило качество томографии большинства мягких тканей, но позволило увидеть сосуды, которые без инъекции контрастного вещества обычно не видны. Удалось показать, что вдоль легкого проходит пара крупных сосудов, которые по своему положению соответствуют легочным артериям и, скорее всего, ими и являются (рис. 12). В жировом органе подобных сосудов нет.

Эти данные, хотя их и нельзя назвать сенсационными, ценны тем, что развеивают последние сомнения насчет того, есть ли у латимерии легкое. Ведь в этом вопросе несколько десятилетий царила путаница. Профессор Смит, например, писал, что «у современного целаканта нет настоящего плавательного пузыря, есть лишь кожаный лоскут — возможно, рудимент некогда существовавшего органа». Почему, собственно, тут идет речь о плавательном пузыре, а не о легком? Да потому что было непонятно, что это вообще такое. Биологи долгое время не были уверены, что перед ними орган дыхания (пусть даже и рудиментарный). Те же Стивен Перри и Маркус Ламбертц еще в 2015 году писали по поводу «странного заполненного жиром непарного органа целакантов», что «остается неясным, имеет ли он отношение к легким» (M. Lambertz, S. F. Perry, 2015. The lung-swimbladder issue: a simple case of homology — or not?). Камила Купелло и ее коллеги наконец прояснили этот момент.
Ископаемых рыб тоже можно исследовать методом компьютерной томографии. И у мезозойского целаканта Macropoma тоже удалось обнаружить отпечатки пары легочных артерий, а заодно и спинной аорты (у «парижской» латимерии спинная аорта была не видна). Здесь все сходится: очевидно, у макропомы легкое снабжалось кровью так же, как и у современной латимерии. В том, что это именно легкое, теперь сомнений не остается. Но вопросы, касающиеся происхождения этого органа, как видим, еще требуют ответов. Впрочем, анатомию кровеносной системы латимерии тоже не помешало бы изучить получше: данные рентгеновской томографии экземпляра, много лет лежащего в формалине, стоит считать скорее предварительными.
Источник: Camila Cupello, Gaël Clément, Marc Herbin, François J. Meunier and Paulo M. Brito. Pulmonary arteries in coelacanths shed light on the vasculature evolution of air-breathing organs in vertebrates // Scientific Reports. 2024. DOI: 10.1038/s41598-024-61065-8.
Сергей Ястребов
Свежие комментарии